Total biovolume of zooplankton collected in upper 50 meters of water column using Hensen nets from R/V Jose Olaya cruises in the Humboldt Current Peruvian coast, 4-18 S, 72-80 W (PCE project)
Project
| Contributors | Affiliation | Role |
|---|---|---|
| Swartzman, Gordon | University of Washington (UW) | Principal Investigator |
| Chandler, Cynthia L. | Woods Hole Oceanographic Institution (WHOI BCO-DMO) | BCO-DMO Data Manager |
Zooplankton were collected using Hensen nets in top 50 meters of the water column from Humboldt Current Ecosystem - Peruvian coast 4-18 degrees South latitude, 72-80 degrees West longitude. Some brief cruise information is available as a PDF file.
Note about data availability:
The sharing of data with other colleagues is subject to an inter-institutional agreement with IMARPE, with some restrictions applying to the use of data by people not directly involved in the agreement, and restrictions for the use of the data after the end of the agreement.
Investigators interested in gaining access to the original data are encouraged to contact:
Patricia Ayòn
IMARPE (Instituto del Mar del Perú)
payon [AT] imarpe [DOT] gob [DOT] pe
Examples from Original Data Files
Original data files exist in Excel spreadsheet files. These include stations (Est), longitude and latitude, date (FECHA), time (HORA), surface temperature (TSM), zooplankton biovolume (Volumen) and numbers of anchovy eggs (Huevos anch) and larvae (Larvas anch).
| Example original data | ||||||||
| Station | Longitude | Latitude | DATE | TIME | Surface Temp | Volume | Eggs | Larvae |
| E-1/C-1JOB | -70.715 | -18.2 | 11/2/2003 | 23:57 | 19.8 | 8 | 411 | 6 |
| E-11/C-2 | -71.176 | -17.907 | 11/3/2003 | 11:33 | 19.26 | 2 | 3 | 39 |
| 14 | -71.363 | -17.724 | 11/3/2003 | 14:15 | 19.15 | 1.5 | 0 | 0 |
Additional processing of these data combined all usable surveys into a single flat (text) file additionally calculated distance to the coast (dc) and to the 200-m isobath (dist200) and added surface temperature (SST) and salinity (SSS) where available. Distances are in km.
| Example processed data | ||||||||
| year | mo | time | lat | dc | dist200 | vol | SST | SSS |
| 1990 | 12 | 1.083333333 | -18.38333333 | 176.192184 | 150.9670611 | 4 | 21.9 | 34.978 |
| 1990 | 12 | 4.083333333 | -18.05 | 107.3411666 | 87.1060464 | 18 | 21.4 | 34.824 |
| 1990 | 12 | 6.733333333 | -17.7 | 20.5228905 | 19.1680014 | 5 | 21.2 | 34.872 |
| 1990 | 12 | 21.35 | -17.48333333 | 3.84008226 | -15.799166 | 10 | 16.2 | 34.734 |
References
Ayón P, and G. Swartzman. (2008) Changes in the long-term distribution of zooplankton in the Humboldt Current Ecosystem off Peru, and its relationship to regime shifts and environmental factors. Fisheries Oceanography, Vol. 17, No. 6. pp. 421-431.
Ayón P, Criales-Hernandez MI, Schwamborn R, Hirche HJ (2008) Zooplankton research off Peru: A review. Prog Oceanogr 79:238-255
(this paper contains references to internal reports describing the data for many of the surveys)
Ayón P, Swartzman G, Bertrand A, Gutiérrez M, Bertrand S (2008) Zooplankton and forage fish species off Peru: Large-scale bottom-up forcing and local-scale depletion. Prog Oceanogr 79:208-214
Ayón, P., Purca, S. and Guevara-Carrasco, R. (2004) Zooplankton volume trends off Peru between 1964 and 2001. ICES J. Mar. Sci Symp. Edition 61(4):478-484
The zooplankton data were collected using a Hensen net drawn vertically through the top 50 meters of the water column. The sampling net had a 0.33 m^2 mouth area with a 300 micrometer mesh cod end. Samples were washed and stored in 2% formalin solution with borax for future processing. Zooplankton biovolume was measured for each sample, as was numbers of anchovy and sardine eggs. The enumeration of species in these samples has barely been started, with occasional attempts at obtaining species composition for given time periods (e.g. 1980-1989).
Additional processing of these data combined all usable surveys into a single flat (text) file additionally calculated distance to the coast (dc) and to the 200-m isobath (dist200) and added surface temperature (SST) and salinity (SSS) where available. Distances are in km.
| Dataset-specific Instrument Name | Hensen Net |
| Generic Instrument Name | Hensen Net |
| Dataset-specific Description | Zooplankton samples were taken with Hensen nets of 0.33 m^2 mouth area with a 300 micrometer mesh, in vertical hauls between 0 and 50 meters. Samples were fixed with 2% formaldehyde buffered with borax. |
| Generic Instrument Description | The Hensen Net was invented by Christian Andreas Victor Hensen (February 10, 1835 - April 5, 1924) who was a German researcher considered by many to be the 'father' of Biological Oceanography. Hensen is credited with first using the term 'plankton', and the Hensen Net is one of several sampling devices he invented to help with sample collection. The Hensen Net has a 'reducing cone' forward of the net mouth, a simple closure system and a cod end collection device.
References:
Hensen, V., 1887. Uber die Bestimmung des Planktons oder des im Meere treibenden Materials an Pflanzen und Thieren. Berichte der Kommssion wissenschaftlichen Untersuchung der deutschen Meere in Kiel 5, pp. 1-107.
Hensen, V., 1895. Methodik der untersuchungen. Ergebnisse der Plankton-Expedition der Humbolt-Stiftung, Lipsius and Tischer, Kiel. |
PCE_multi
| Website | |
| Platform | R/V Jose Olaya |
| Start Date | 1963-01-01 |
| End Date | 2006-12-31 |
| Description | Fishing catch and effort data were collected aboard the 800 purse seiner strong Peruvian industrial fleet. A program of observers at sea on board fishing vessels started in 1996 and provides, on a daily basis, information about the fishing trips (boat reference, hold capacity, trip dates, hailing port, travel duration, searching duration, total catch, number of fishing sets) and the fishing sets (exact spatial position, catch, species composition) of about 25 purse-seiners randomly selected and distributed all along the Peruvian coast.
Multiple vessels were involved including the R/V Jose Olaya operated by IMARPE (Instituto del Mar del Peru).
No cruise metadata was contributed with the results from this project. Cruise track location data were entered in the database to describe roughly the cruise study area off the coast of Peru (positions and times reflect the study area in time and space, but not a cruise track). The data from these cruises are not expected to be contributed to BCO-DMO and will not be archived at a US National Data Center.
The data are available from a Peruvian data repository. Please refer to the project description for contact information. |
Biological response to regime shifts and ENSO events in the Peruvian Humboldt Current ecosystem (PCE)
The primary aim of this project was to examine the long-term patterns in distribution and abundance in the HCE of the two major pelagic fish species, Anchovy (Engraulis ringens), and Sardine (Sardinops sagax). We will determine the relationship of these patterns to environmental factors (e.g. currents, temperature, bathymetry, eddy vorticity) and to the distribution and abundance of the primary prey resources of anchovy and sardine: copepod, euphausiid and other dominant zooplankton species. To this end we tested hypotheses concerning the spatial aggregation and overlap of these species, expected shifts in fish and zooplankton abundance and biomass distribution during warm and cold regimes (and the extreme temperature anomaly periods of El Niño and La Niña), and the relationship of fish stomach fullness and diet to prey availability.
The project was done in collaboration with researchers at IRD (Institut de Recherche pour le Développement, France) 'ECO-UP' UR 097 and of the Interdepartmental Thematic Action "Humboldt Current System" and CENSOR (EU project run from University of Kiel, Germany). These people were acknowledged as co-investigators, but no contact information was given: Arnaud Bertrand, Sophie Bertrand (postdoctoral associate), Pierre Fréon and François Gerlotto IRD (Institut de Recherche pour le Développement, France) and Renato Guevara, Patricia Ayòn and Mariano Gutiérrez IMARPE (Instituto del Mar del Perú).
Data Sources
The primary sources of data for this project are the pelagic fish acoustic surveys by IMARPE (Instituto del Mar del Peru) conducted for abundance estimation and distribution of pelagic and demersal fish resources (Figure 1) and zooplankton and phytoplankton net surveys conducted over the same area (many in conjunction with acoustic surveys) since 1960. Since 1998 the acoustic data have been collected in digital form, using a SIMRAD hull-mounted split beam EK-500 echo sounder system at 38 and 120 kHz. Before 1998 a series of other SIMRAD echo sounder system were used in accordance with the available technology at the time. Abundance estimates for the major commercially exploited pelagic fish species (sardine and anchovy, horse mackerel and mackerel), based on these acoustic data and accompanying trawl data, were recorded from 1983-2003 as well as some schooling characteristics. All these data are in digital form. At least two acoustic surveys were run in most years. Extensive midwater trawl surveys accompanied the acoustic surveys for species identification with an average of two trawls per transect for the entire survey (Figure 1). The seasonal and temporal distribution of scientific acoustic surveys is: Spring (1983, 1986-87, 1989, 1997-2003 [twice in 1999 and 2002]), summer (1990-96, 1998-2003), autumn (1985-86, 1997-98), and winter (1984, 1988-89, 1991, 1996, 1998-2001). During acoustic surveys sea surface temperature, salinity, oxygen, nutrients and chlorophyll were collected and underway observations of birds and mammals were recorded since 1998. CTD (Conductivity-Temperature-Depth) and XBT (expendable bathythermogram) drops were performed at trawl locations and provide information on temperature profiles, thermocline depth and geostrophic flow over the survey area.
Since 1996, anchovy stomachs were collected from random samples of one, two or three fish per size class (8-9.5, 10-11.5, 12-13.5, 14-15.5, 16-17.5, 18-19.5 mm) from each trawl. The fish length, weight and sex were recorded from each sample, which were preserved in a 5% formaldehyde-water mixture for later analysis in the laboratory. Individual stomach contents were weighed to the nearest 0.001 g and used to calculate stomach fullness. The stomach contents were identified to genera level and numbers of each diet component were counted. Reproductive condition samples of anchovy were also taken from each trawl. The percentages of each reproductive stage were summarized, and Stage 4, which scored for the presence of post ovulatory follicles, was used to calculate spawning fraction and assess reproductive condition.
Fishing catch and effort data are available from the 800 purse seiner strong Peruvian industrial fleet. A program of observers at sea onboard fishing vessels started in 1996 and provides, on a daily basis, information about the fishing trips (boat reference, hold capacity, trip dates, hailing port, travel duration, searching duration, total catch, number of fishing sets) and the fishing sets (exact spatial position, catch, species composition) of about 25 purse-seiners randomly selected and distributed all along the Peruvian coast. Since 1998, a VMS (SISESAT: SIstema de SEguimiento SATelital) has been operational for the entire Peruvian industrial fishing fleet. Each vessel is equipped with an Argos satellite transmitter (CLS Argos, Toulouse, France) furnished with its own Global Positioning System (GPS) receiver. Each transmitter sends its time-referenced position with an accuracy of 100 m on an hourly basis. These data are important because they are spatially and temporally extensive and fill in time periods between the scientific surveys. Fleet sampling is also a valuable source of additional anchovy stomach content and reproductive condition data, which have been collected since 1998 using the same protocols as for the survey data.
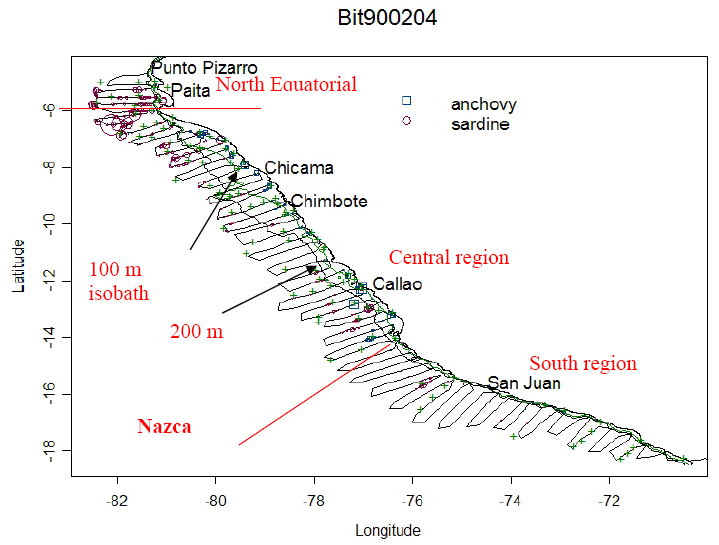
Figure 1. Example of distribution of four species of pelagic fish from an acoustic survey performed during February-April 1990 by IMARPE. Transect lines are shown by spaced dots. This survey was during a period of sardine dominance. Data are summarized on an ESDU (Elementary Sampling Distance Unit, 1 n. mi) basis along the transects. Areas having no circles had no fish of these species. Also shown are the locations of zooplankton net and trawl samples for this surve y (green x). The locations of major ports are shown where IMARPE maintains oceanographic observation transects. The figure also shows oceanographically and bathymetrically defined region boundaries within Peru for the HCE (note differences in the 100 and 200 m isobaths along the coast). The Nazca Ridge defines the boundary between the central and southern HCE regions.
Zooplankton and phytoplankton net samples have been collected on all acoustic surveys at the trawl locations (Figure 1) as well as during many other surveys, beginning in 1961. The zooplankton data were collected using a Hansen net drawn vertically through the top 50 meters of the water column with a 300 micrometer mesh cod end. Samples were washed and stored in 2% formalin solution with borax for future processing. Zooplankton biovolume was measured for each sample, as was numbers of anchovy and sardine eggs. The enumeration of species in these samples has barely been started, with occasional attempts at obtaining species composition for given time periods (e.g. 1980-1989). The phytoplankton data were collected using a 5 minute red standard surface tow with a 75 micrometer net mesh. These data have all been enumerated to the species level and were recorded at two levels of resolution; 1) biovolume and qualitative enumeration of dominant species and 2) exact counts and species enumeration. All surveys have been enumerated at the lower resolution, with data from selected surveys in the 1980's still unprocessed at the higher resolution. These data exist mostly in paper form, except 1961-1965, which were entered into EXCEL spreadsheets as part of an agreement with the World Oceanographic Data Center (Sydney Levitus, Director, Silver Spring MD).
Data Processing - pelagic fish
The assignment of acoustic backscatter to fish species is carried out by 'scrutinizing' the data, which depends on correlation with the accompanying midwater trawls, which are conducted frequently along the survey transects (See Figure 1), and pattern recognition of the echogram shapes and locations of pelagic fish schools. Scrutinizing was done by IMARPE scientists (for data since 1998) after each survey using Echoview (Sonardata Pty. Ltd) a software package which provides a high-resolution echogram (0.25 to 1 m vertical x approximately 2 m horizontal depending on ping rate and boat speed) at both operating frequencies (38 and 120 kHz) and image processing tools to manually classify fish school backscatter. Data before 1998 were more tediously processed from paper echograms and cumulative acoustic backscatter (sA) information at both frequencies. Models of anchovy and sardine target strength (backscatter as a function of size) were used, along with fish size distributions from the trawls to transform acoustic backscatter (sA) to biomass. Scrutinizing was simplified by sardine and anchovy almost always occurring in single species school aggregations (Mariano Gutierrez, IMARPE, personal communication). As well, IMARPE conducts accompanying trawls more than twice as frequently as most other fisheries stock assessment survey teams (Figure 1), which provides assurance of the quality of the data.
Zooplankton size and species composition
Zooplankton size distribution were qualitatively ascertained for all surveys from 1961 to the present.
Hypotheses concerning HCE functional processes related to climatic variability
We organized the work around a set of hypotheses concerning changes in distribution and abundance of fish and zooplankton to regime-shift and ENSO changes in the HCE and predator-prey dynamics (Exhibit 1).
Exhibit 1. Hypotheses concerning the effect of regime shifts on the distribution and abundance and feeding of biota at different trophic levels. Hypotheses are followed by rationale (R), evidence (E), data sources (D), statistical methods used (S), sample unit (U) and comments (C).
Zooplankton size composition and distribution
1. Over the Central Peruvian coast to a distance of approximately 100 n.mi abundance of smaller zooplankton is higher offshore, while nearshore upwelling areas are dominated by larger zooplankton.
R. Cooler waters nearshore should favor slower growing and later maturing but large zooplankton, while warmer waters further offshore should favor faster growing but smaller zooplankton because colder waters induce metabolic slowing but are nutritionally richer as a result of deeper upwelling.
E. Anchovy, which are closer to shore than sardine, favor larger copepods and euphausiids (Pepe Espinosa, IMARPE, unpublished data). Sardine, primarily filter feeding, are limited in this feeding mode to zooplankton smaller than 1.2 mm. There was a shift in abundance of zooplankton toward smaller zooplankton during the warm El Nino conditions of 1997-98. However, others found a shift between larger copepods offshore to smaller copepods nearer shore in a transect off Chimbote (Figure 1).
D. Zooplankton biovolume from net samples 1961-2004; qualitative by-eye estimates for zooplankton size distribution. Survey temperature and salinity data to define water masses (since 1983) Water mass reconstructions from before 1983.
S. ANOVA; PCA[1] to explore overall relations between factors; CART[2] on zooplankton size as a function of latitude, distance to coast, water mass (Temperature-Salinity)
U. Locations of zooplankton net hauls
C. In addition to distance from coast, water mass and latitude will be used as covariates to more fully examine changes in size distribution.
2. Euphausiids should extend further south during El Niño and El Viejo than during La Nina and La Vieja periods
R . The strengthened Cromwell undercurrent combined with diel migration of euphausiids should transport them further south
E. Swartzman and Hickey (2003) and Swartzman et al (2005) found northward shifts in euphausiid abundance in summer 1998 compared to 1995, 1998 being an El Niño year, in the California Current Ecosystem. Blascovic' et al., (1999) found euphausiid abundance in anchovy stomachs to be higher in the southern region of the Peruvian HCE during the 1997-98 El Niño.
D. Zooplankton net hauls 1961-2003 processed with ZOOSCAN to locate those in euphausiid size range (10-25 mm). Anchovy stomach content data 1996-2003.
S. GAM [3] on euphausiid abundance according to latitude and distance from coast at each period; t-tests on centroid of euphausiid abundance and of percentage euphausiids in anchovy stomachs in north, central and south regions for cool and warm regime periods.
U. Individual net hauls; trawl hauls; Industrial fishery collection from individual purse seine net sets.
C. Species composition may only be available for the transect off Chimbote, but size distribution can be used to 'identify' euphausiids on other transects.
3. The species composition of zooplankton off Peru should shift towards more equatorial species during El Niño and El Viejo and towards more austral species during La Niña and La Vieja periods.
R. Stronger surface winds and upwelling should create cooler conditions during La Niña and transport austral zooplankton north, whereas during El Niño diel migrating equatorial zooplankton will be transported southward by the strengthened Cromwell current.
E. The dominant medium sized copepod, Calanus chilensis, was abundant in northern Chile during the 1997-1998 El Niño (Gonzalez et al 2000, Ulloa et al., 2001) though it was restricted to near-shore sites. Zooplankton assemblages shifted to smaller species during this event but the total biomass did not change (Escribano et al., 2004). Santander and Carrasco (1985), Carrasco and Santander (1987) and Abanto (2001) found increased abundance of large carnivorous copepods and reduction of herbivorous copepods during the 1983 El Niño. However, the origin of these species (i.e. austral, temperate or equatorial) has not been checked. Shifts in abundance from more temperate to more boreal species were found after the regime shift of 1997-1998 off California, Oregon and British Columbia in the CCE by Ohmann (2003), Peterson and Mackas (2001) and Mackas et al. (2001) respectively.
D. Zooplankton species composition for a single transect off Chimbote 1964-2004; temperature anomaly data off Chimbote (Figure 1).
S. c2 and ANOVA on percent temperate species along chosen transects through 3 regimes (cold-warm-cold) since 1964. Factorial correspondence analysis [4] on zooplankton types with environmental factors as illustrative parameters projected on factorial space.
U. Zooplankton net hauls, genera composition on a single transect using ZOOSCAN images
Pelagic Fish
4. Cooler regimes should favor anchovy, which spread further offshore during these conditions. Sardine should move further offshore or further north and be less abundant off Peru. The sub-Antarctic species Munida (Pleuroncodes monodom) expands further north into Peru.
R. The distribution of pelagic fish follows the range of their ecological niche (water mass characteristics, plankton size, etc.). Anchovy distributions are strongly related to cold coastal water that expand offshore during cold periods (Bertrand et al., 2004b; Gutierrez et al., 1999). During a cooler regime and La Niña, the sardine, as observed in the CCE by Rodriguez-Sanchez et al. (2002), may move further north, out of the HCE. Stronger Humboldt Current water transport that prevails near the coast in Southern Peru during La Vieja allows expansion into Peru of sub-Antarctic species (e.g. Munida).
E. The pelagic fish in the HCE are dominated by anchovy and sardine which, as in the CCE, appear to oscillate out of phase (Lluch-Belda et al., 1989). Acoustic surveys during the La Niña, which occurred within the cold decadal regime (e.g. in summer 1999), showed dominance of the pelagic fish by anchovy (Gutierrez et al., 1999). Bertrand et al., (2004b) found that anchovy abundance appeared lower during the El Niño, but suggested that fish were probably aggregated inshore of the study transects. Sardine also moved near shore. During the La Niña of 1998-99 sardine dispersed offshore. Munida appeared off southern Peru after 1995 and abundances have remained high except for the El Niño period (Mariano Gutierrez, IMARPE, unpublished data).
D. Acoustic survey and trawl data from 1983-2004.
S. GAM on abundance as a function of latitude, distance to the coast and other environmental parameters at each period; CART of centroid of distance from coast for sardine and anchovy as function of year, temperature anomaly, season. t-tests for Munida abundance during warm and cooler years.
U. Data unit can be either ESDU (1-2 n.mi.) or individual schools. Acoustic data will be combined with targeted fishing trawls.
C. Species composition from hauls and scrutinizing will be evaluated using the AICASA method.
5. Anchovy abundance should predominate in cooler upwelling water or mixed upwelling-oceanic water. Sardine should be found primarily in oceanic water or mixed upwelling water. Anchovy and sardine generally shift in distribution seasonally, aggregating near shore during summer when upwelling is more shoreward and dispersing offshore during the other seasons of more pervasive offshore upwelling. Aggregation during summer is stronger during a warm regime (and El Niño) than during a cool regime (and La Niña).
R. Anchovy prefers upwelling water, larger zooplankton and diatoms, while sardine prefer and more efficiently utilizes small zooplankton such as small diatoms and flagellates, which predominate further offshore and in warm conditions (Escribano et al., 2004). During summer, upwelling is often confined to coastal waters with oceanic waters just offshore. During the other seasons an offshore extension of coastal upwelling conditions allows dispersal of anchovy (M. Gutierrez, IMARPE, unpublished data).
E. Bertrand et al. (2004b) found these sardine and anchovy water mass associations for surveys during the El Niño - La Niña period of 1997-1999, with high aggregation of sardine and anchovy onshore during the 1997-1998 El Niño. Mariano Gutierrez (IMARPE, personal communication) developed this hypothesis suggested by acoustic survey results and the high CPUE of the anchovy fishery during summer months, especially during warmer regimes, due to concentration of anchovy near shore. See Figure 1 for an example of summer anchovy distribution near shore.
D. Acoustic survey data 1983-present. Temperature and salinity data from surveys. Uses an algorithm for water mass definition (Luis Vasquez, IMARPE, personal communication).
S. Canonical correlation [5] of abundance of sardine and anchovy and of water types. GAM of sardine and anchovy presence-absence and water types using a binomial probability model.
U. ESDU level acoustic data on sardine and anchovy as in hypothesis 4) above.
6.Anchovy abundance may be highest along the interfaces between strong counterclockwise and clockwise eddies.
R. These are regions of the greatest mixing, providing higher nutrients and further offshore than the shelf region. The longevity of these features (1-2 months; Hormazabal et al 2004) suggests they can provide stable feeding areas for anchovy.
E. Fernando Espindola (Univ of Concepcion, unpublished manuscript) found the highest abundance of jack mackerel, a strong predator on anchovy, in the current interfaces between counterclockwise and clockwise eddies off central Chile (30 degrees S) during 1997-1998. Front features appear to serve as aggregation areas for anchovy schools (M. Gutierrez, IMARPE and F. Gerlotto, IRD, unpublished results of a 2004 IMARPE cruise directed at studying anchovy schooling behavior).
D. Acoustic survey data 1995-2004. ERS-1 and 2 and TOPEX/Poseidon data on sea surface altitude. Definition of eddies using methods of Hormazabal et al (2004).
S. F tests for correlation of abundance with eddy vorticity. GAM of abundance as a function of latitude, coast distance, and eddy vorticity (F. Espindola, unpublished manuscript).
U. Acoustic data by ESDU. Also possible to use school clusters [6] as the sampling unit.
C. Eddy vorticity will be interpolated [7] onto acoustic ESDU locations or cluster centroid locations.
Predation and feeding
7. Anchovy have higher stomach fullness during periods of higher macrozooplankton abundance
R. Higher food abundance should result in better feeding conditions and fuller stomachs
E. P. Espinoza and B. Buitron (IMARPE, unpublished data) examined anchovy stomach contents and fullness from 1999 to 2003 and found stomach fullness to be higher in successful recruitment years
D. Anchovy stomach data 1996 - 2004. Zooplankton net data from the same surveys at the same locations. Also additional zooplankton surveys to determine the period of high zooplankton abundance and Industrial fishery collected stomachs will be used.
S. t-tests on stomach fullness for high and lower periods of macro zooplankton abundance.
U. Location of trawl hauls and purse seine catches.
C. High and low zooplankton abundance ranges will be defined by percentiles of abundance for all available surveys.
Approach and Analysis Methods
Data Fusion and Analysis
Testing the proposed hypotheses required combining long-term acoustic survey data with environmental and biological data. The melding of these data is possible because they were simultaneously collected and all are geo-referenced and time-stamped. To allow comparison of changes in species composition and distribution over time we stratify the data into oceanographically defined regions (north-south) and zones (onshore-offshore). We define region boundaries (Figure 1) based on long-term sea surface temperature trends and on bathymetric differences. An equatorial North region, north of Punta Falsa, near Paita, Peru, has a shelf-break close to the coast. The Central region from Punta Falsa to the Nazca Ridge area south of Pisco, Peru is dominated by the Humboldt (north flowing surface water) and Cromwell (south flowing deep) currents and features a wide shelf. The South region extends south to about 30oS. It has a shelf break near to the coast and upwelling tends to be more coastal and seasonal than further north. Additionally, we will separate the regions into zones based on bathymetry, including the shelf (bottom depth < 200 m), shelf-break (200-800 m) and coastal offshore (> 800 m.). The choice of fixed zones and regions for an area as labile as the HCE is a pragmatic expedient. An alternative approach is to define regions by water masses. Initial subdivision of the survey area into water masses defined by their ranges of surface temperature and salinity (Luis Vasquez, IMARPE, personal communication) resulted in four primary water mass types, (cold upwelling water, offshore water, tropical water and equatorial water) and four mixed water masses, which accounted for the entire survey area in almost all cases (that is all surface measurements fell uniquely into one of the water mass types. We used both the water mass modality to define regions and the fixed boundary regions and bathymetric zones depending on the question asked.
Longer term regime shift and seasonal patterns in the HCE were included using monthly temperature anomalies from long-term observation stations off the Peruvian coast near Paita, Chicama, Chimbote, Callao and San Juan de Marcona (Figure 2).
Relevant publications by PI and collaborators:
Ayòn P, & G. Swartzman. (2008) Changes in the long-term distribution of zooplankton in the Humboldt Current Ecosystem off Peru, and its relationship to regime shifts and environmental factors. Fisheries Oceanography, Vol. 17, No. 6. pp. 421-431.
Ayòn P, Criales-Hernandez MI, Schwamborn R, Hirche HJ (2008) Zooplankton research off Peru: A review. Prog Oceanogr 79:238-255
Ayòn P, Swartzman G, Bertrand A, Gutiérrez M, Bertrand S (2008) Zooplankton and forage fish species off Peru: Large-scale bottom-up forcing and local-scale depletion. Prog Oceanogr 79:208-214
Ballòn, M., Wosnitza-Mendo, C., Guevara-Carrasco, R., Bertrand, A (2008) The impact of overfishing and El Niño on the condition factor and reproductive success of Peruvian hake, Merluccius gayi peruanus. Prog Oceanogr 79:300-307 (Science Direct)
Bertrand A. Segura M. Gutiérrez M. Vásquez L. (2004) From small-scale habitat loopholes to decadal cycles: a habitat-based hypothesis explaining fluctuation in pelagic fish populations off Peru. Fish Fish 5:296-316
Bertrand A., Gerlotto F., Bertrand S., Gutiérrez M., Alza L., Chipollini A., Diaz E., Espinoza P., Ledesma L., Quesquén R., Peraltilla,S., and Chavez F. (2008). Schooling behaviour and environmental forcing in relation to anchoveta distribution: An analysis across multiple spatial scales. Progress in Oceanography, 79: 264-277.
Bertrand S., Dewitte B., Tam J. Diaz, E. & Bertrand A. (2008). Impacts of Kelvin wave forcing in the Peru Humboldt Current system: Scenarios of spatial reorganizations from physics to fishers. Progress in Oceanography, 79:278-289.
Bertrand S., Burgos J. M., Gerlotto F. & Atiquipa J. (2005). Lévy trajectories of fishers as an indicator of pelagic fish spatial distribution: the case of the Peruvian anchovy (Engraulis ringens) fishery. ICES Journal of Marine Science, 62: 477-482.
Bertrand, S., Díaz, E. & Lengaigne, M. (2008). Patterns in the spatial distribution of Peruvian anchovy (Engraulis ringens) revealed by spatially explicit fishing data. Progress in Oceanography, 79: 379-389.
Bertrand S., Bertrand A., Guevara-Carrasco R. & Gerlotto F. (2007). Scale invariant movements of fishermen: the same foraging strategy as natural predators. Ecological Applications, 17(2):331-337.
Brochier, T., Lett, C., Tam, J., Fréon, P., Colas, F., Ayòn, P. (2008) An individual-based model study of anchovy early life history in the northern Humboldt Current system. Prog Oceanogr 79:313-325 (Science Direct)
Chavez, F., Bertrand, A., Guevara-Carrasco, R., Soler, P., Csirke, J. (2008) The northern Humboldt Current System: Brief history, present status and a view towards the future Prog Oceanogr 79: 95-105 (Science Direct)
Espinoza P, Bertrand A (2008) Revisiting Peruvian anchovy (Engraulis ringens) trophodynamics provides a new vision of the Humboldt Current system. Prog Oceanogr 79: 215-227.
Espinoza P, Bertrand A, van der Lingen CD, Garrido S, Rojas de Mendiola B (2009) Diet of sardine (Sardinops sagax) in the northern Humboldt and comparison with the diets of clupeoids in this and other eastern boundary upwelling systems. Prog Oceanogr 83:000-000
Estrella, C. & G. Swartzman (2009) The Peruvian Artisanal Fishery: Changes in patterns and distribution over time. Fisheries Research (in press)
Gerlotto F, Bertrand S., Bez N. & Gutiérrez M. (2006). Waves of agitation inside anchovy schools: a way to transmit information and facilitate fast morphological and structural changes in response to predation, as observed with multibeam sonar. ICES Journal of Marine Science, 63:1405-1417
Gutiérrez M., Swartzman G., Bertrand A. & Bertrand S. (2007). Anchovy and sardine spatial dynamics and aggregation patterns in the Humboldt Current ecosystem, Peru, from 1983-2003. Fisheries Oceanography, 16(2): 155-168.
Gutiérrez M., Ramirez A., Bertrand S., Moròn O., & Bertrand A. (2008). Ecological niches and areas of overlap of the squat lobster 'munida' (Pleuroncodes monodon) and anchoveta (Engraulis ringens) off Peru. Progress in Oceanography, 79:256-263.
Lett C, Penven P, Ayòn P, Fréon P (2007) Quantification of enrichment, concentration and retention processes in relation to anchovy (Engraulis ringens) ichthyoplankton distribution in the northern Humboldt upwelling ecosystem. J Mar Sys 64:189-200.
Paramo J., Bertrand S., Villalobos H. & Gerlotto F. (2007). A three-dimensional approach to school typology using vertical scanning multibeam sonar. Fisheries Research, 84:171-179.
Simmonds J, Gutiérrez M, Chipollini A, Gerlotto F, Woillez M, Bertrand A (2009) Optimizing the design of acoustic surveys of Peruvian anchoveta. ICES J Mar Sci 66:1341-1348
Swartzman G., Bertrand A., Gutiérrez M., Bertrand S. & Vasquez L. (2008). The relationship of anchovy and sardine to water masses in the Peruvian Humboldt Current System from 1983 to 2005. Progress in Oceanography, 79:228-237.
Tremblay Y., Bertrand S., Henry R.W., Kappes M. A., Costa D. P., Schaffer S. A. (2009). Analytical approaches to investigate seabird-environment interactions: a review. Marine Ecology Progress Series (in press).
Tam, J., Taylor, M.H., Blaskovic, V., Espinoza,P., Ballòn, M., Díaz, E., Wosnitza-Mendo, C., Argüelles, J., Purca, S., Ayòn, P., Quipuzcoa, L., Gutiérrez, D., Goya, E., Ochoa, N., Wolff M. (2008) Trophic modeling of the Northern Humboldt Current Ecosystem, Part I: Comparing trophic linkages under La Niña and El Niño conditions. Progress in Oceanogr, 79:352-365 (Science Direct)
Taylor, M.H., Tam, J., Blaskovic, V., Espinoza, P., Ballòn, M., Wosnitza-Mendo, C., Argüelles, J., Díaz, E., Purca, S., Ochoa, N., Ayòn,P., Goya, E., Gutiérrez,D., Quipuzcoa, L., Wolff, M. (2008) Trophic modeling of the Northern Humboldt Current Ecosystem, Part II: Elucidating ecosystem dynamics from 1995 to 2004 with a focus on the impact of ENSO. Progress in Oceanogr, 79:366-378 (Science Direct)
van der Lingen CD, Bertrand A, Bode A, Brodeur R, Cubillos L, Espinoza P, Friedland K, Garrido S, Irigoien X, Möllmann C, Rodriguez-Sanchez R, Tanaka H, Temming A (2009) Chapter 8, Trophic dynamics of small pelagic fish. In: Checkley DM Jr, Roy C, Alheit J, Oozeki Y (Eds) Climate Change and Small Pelagic Fish. Cambridge University Press, 112-157.
Footnotes:
[1] PCA (Principal Components Analysis) is a statistical tool for transforming linear combinations of covariates into a smaller number of orthogonal components which maximize the variance they explain of the original data.
[2] CART (Classification and Regression Trees) is a model-based tree clustering algorithm. A model relationship is set up between a nominal (Regression) or categorical (Classification) dependent variable and concomitant covariates. CART finds a series of dichotomous cuts (branches) of covariates that provide the minimum deviance (for regression) or rate of incorrect classification (for classification) for both groups of data (separated based on some threshold of one of the covariates) and continues to split the data in tree fashion until some minimal number of data points remain in each class. While CART does not provide a statistical test for significance, it is an excellent tool for uncovering relationships between a complex set of variables in a large data set. Thus, CART may be used to decide which covariates provide the best prediction for fish or zooplankton abundance or species composition. A major advantage of CART for our data is that it can handle categorical dependent variables as well as categorical covariates and makes no assumptions about the underlying probability distributions.
[3] GAM (Generalized Additive Models, Hastie and Tibshirani 1990) is a non-parametric smoothing-based regression method that relaxes the assumption of linearity between the dependent variable (e.g. anchovy abundance per ESDU) and the covariates (e.g. temperature, salinity, oxygen, bottom depth). GAM allows the underlying probability distribution of the dependent variable to be assumed from any distribution in the exponential family, including the normal, Poisson, binomial and gamma distributions. Asymptotic significance tests (either F or c2) are calculated for each covariate. Interactions between covariates are allowed. Since GAM solutions are not guaranteed to converge, a penalty is paid for having extraneous covariates. CART can be used to first identify the most important covariates.
[4] An exploratory method for looking at relationships between multiple categorical variables using data normalization, canonical eigenanalysis and a distance measure for similarity (Lebart et al., 2000).
[5] Canonical correlation is a multivariate statistical technique for relating two independent sets of data using the correlation between important variables in each data set (Lebart et al., 2000).
[6] School information is obtained directly from the high resolution acoustic data. School clusters are defined as groups of schools within a threshold distance of their next neighbors. As such, they may span several or parts of several ESDU. They have the advantage of not being tied to the ESDU resolution (1-2 n.mi.).
[7] Interpolation will be done using kriging methods, which include spatial autocorrelation and trends (R functions geoR and geoRglm).
| Funding Source | Award |
|---|---|
| NSF Division of Ocean Sciences (NSF OCE) |
[ table of contents | back to top ]