Cell counts from phytoplankton Si utilization experiments during 8-day laboratory cultures in 2016 and 2017
Project
| Contributors | Affiliation | Role |
|---|---|---|
| Taylor, Alison | University of North Carolina - Wilmington (UNC-Wilmington) | Principal Investigator |
| Meyer, Erin Michelle | University of North Carolina - Wilmington (UNC-Wilmington) | Student |
| York, Amber D. | Woods Hole Oceanographic Institution (WHOI BCO-DMO) | BCO-DMO Data Manager |
Abstract
The methodology below describes all data collected from this experiment. This dataset landing page serves the "Si Depletion Experiment: Cell Counts" data. Other datasets can be found in the "Related Datasets" section of this page.
Experiment Overview: Phytoplankton Si utilization during an 8 d culture experiment was assessed. A variety of silicifying and non-silicifying species were grown in fully amended LH and F/2 media supplemented with 5 M Si. All species were grown in quadruplicates except for Thalassiosira weissflogii and the no cells control. Aliquots of cultures for cell counts, dissolved Si (DSi), and biogenic Si incorporated into the cells (BSi) were taken daily from T0 – T4, then on T6 and T8 and subsequently processed as described below. In addition, EDS analysis was used to determine if Si was present in biomineral structures.
Cell Counting: Cells were counted on each collection day using a hemocytometer or Sedgwick-Rafter chamber. A minimum of 300 cells were counted per sample. Growth curves were plotted and specific growth rates were calculated for each species throughout the 8 d sampling period.
DSi Method: Autoclaved and filtered Gulf Stream seawater was amended with LH or F/2 nutrients and [Si] measured prior to adding sufficient NaSiO3 to reach a starting [Si] of 5 M. Cells were harvested from cultures at early exponential phase and gently washed in Si-free media using a 0.4 m Nalgene polycarbonate filter unit. Replicate 200 mL cultures were seeded with washed cells for a starting density of 1-5 x 104 mL-1. Each sampling day, 15 mL culture aliquots were 0.2 m filtered (Merck Millipore Ltd.) and filtrate was stored at 4C prior to DSi analysis. The filters were frozen for later BSi analysis (see below). For AutoAnalyzer nutrient analysis, the molybdate method was used (modified from Brzezinski and Nelson (1995) and Brzezinski et al. (1997)). The system was washed with sodium dodecyl sulfate (SDS) to lubricate the Si lines. Oxalic acid concentrations were increased to saturated levels (143 g/L) to overcome any phosphate interference. Molybdate was made fresh for each run. The tubing diameter for the oxalic line was also increased to 0.035 in, with a flow rate of 0.41 at 40%.
BSi Method: The filters (See DSi explanation above) with cells were frozen at -20C prior to BSi analysis. For BSi determination, silicifying species were collected on 0.2 m polycarbonate filters, and processed using the alkaline digestion method as described by Brazinski and Nelson (1995), with modifications from Paasche (1973) and Krausse (1983). For coccolithophores, filters were first treated with 1mL 0.5 M HCl to fully dissolve coccoliths (Moheimani & Borowitzka, 2006), then treated with 4 mL 0.2 M NaOH to neutralize, before the alkaline digestion. For the alkaline digestion, each filter was placed in a 15 mL polymethylpentene tube (Diagenode, Inc.) with 4 mL of 0.2 M NaOH and brought to 100oC in a water bath for 20 min, cooled, and neutralized with 1 mL of 0.5 M HCl. The digest was centrifuged at 10,000 rpm for 9 minutes and aliquots of supernatant were removed and diluted for autoanalyzer analysis as appropriate. Analytical blanks with filter only (no cells) were included for each run. Autoanalyzer conditions are the same as described under the DSi Collection section.
EDS Method: For EDS analysis 1-3 mL of culture were filtered onto 13 mm 0.4 m isopore filters [Merck Millipore Ltd.] and rinsed with Nanopure water buffered to pH 8.0 with 1 mM HEPES to remove salts. Filters were air-dried and mounted onto a SEM stub with carbon adhesive tabs before coating with 10 nm Pt/Pd. EDS analysis was performed at the Joint School of Nanoscience and Nanoengineering (Zeiss Auriga SEM, with a Bruker Quantax detector and analysis software) or at North Carolina State University (FEI Verios 460L SEM, with an Oxford Xmax silicon drift EDS detector and AZtec acquisition and analysis software). A minimum of 500,000 counts were collected, between 2,000 – 8,000 cps with an average deadtime < 5%. Standardless quantification was used to determine atomic % and weight % for each element.
Cell Counts:
The formulas for growth rate and doublings per day were captured as the image "cell_counts_processing.png" which is displayed below and can also be found in the "Supplemental Files" section.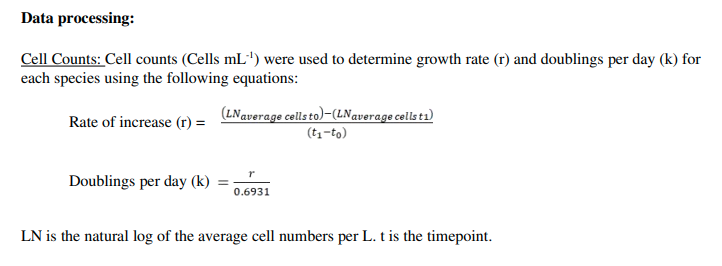
BCO-DMO data manager processing notes:
* Scientific names were checked using the World Register of Marine Species "Taxa Match" Tool. All names matched accepted names exactly from file "Si Depletion Experiment Dataset.xlsx" sheet name "EDS" column "Species." Species list with the AphiaIDs added as a supplemental file.
* Data for this Cell counts dataset imported into the BCO-DMO data system from file "Si Depletion Experiment Dataset.xlsx" sheet name "Cell_Counts"
| File |
|---|
cell_counts.csv (Comma Separated Values (.csv), 6.58 KB) MD5:7784fa104a0f30721d406f74536a2b9f Primary data file for dataset ID 858823 |
| File |
|---|
Data processing formulas for "Si Depletion Experiment: Cell Counts" filename: cell_counts_processing.png (Portable Network Graphics (.png), 42.70 KB) MD5:a856a4febec58634355419f209c06ce9 Image showing the data processing formulas. |
Species list filename: species_list.csv (Comma Separated Values (.csv), 235 bytes) MD5:15648a45a0bd7a90c92e054ad3645135 Species list and AphiaIDs for species in Si depletion experiments. More information about AphiaIDs can be found at the World Register of Marine Species (WoRMS). |
Relationship Description: Data from the same experiment.
Relationship Description: Data from the same experiment.
Relationship Description: Data from the same experiment.
| Parameter | Description | Units |
| Sample | The sample of cells that were being counted. Name is in the following order: Species_Replicate flask number. The species used were Prymnesium neolepis, Thalassiosira weissflogii, Emiliania huxleyi, Gephyrocapsa oceanica, Calcidisdcus leptoporus, Coccolithus braarudii, and Scyphosphaera apsteinii. | unitless |
| Sampling_Day | Day aliquots were taken from flasks and cells were counted. Numeric day of the experiment for the sample starting at day 0. | days (d) |
| Cells_per_mL | The amount of cells of each species in 1 mL of growth media on the day of sampling. Cells were counted with either a Sedgwick-Rafter or Hemocytometer. | Cells per milliliter (Cells/mL) |
| Dataset-specific Instrument Name | |
| Generic Instrument Name | Microscope - Optical |
| Dataset-specific Description | light microscope with either a Hemocytometer or Sedgewick-Rafter chamber |
| Generic Instrument Description | Instruments that generate enlarged images of samples using the phenomena of reflection and absorption of visible light. Includes conventional and inverted instruments. Also called a "light microscope". |
NSFGEO-NERC: An unexpected requirement for silicon in coccolithophore calcification: physiological, ecological and evolutionary implications (Coccolithophore Silicon Requirements)
NSF Award Abstract:
Biomineralization by marine phytoplankton has had a profound impact on our planet. The production of special cell wall material, calcite coccoliths by coccolithophores and silica frustules by diatoms, are major drivers in global biogeochemical cycles, but the underlying cellular processes remain poorly understood. It is widely considered that calcification in coccolithophores occurs through a very different process to silicification in diatoms, however some ecologically important coccolithophore lineages possess diatom-like silicon (Si) transport systems and have an absolute requirement for Si during coccolith formation. Importantly, the abundant bloom-forming coccolithophores such as Emiliania huxleyi exhibit no requirement for Si. There is a clear need to understand how these different physiological requirements for dissolved Si have driven the ecology and evolution of the coccolithophores. The project will yield a more complete understanding of the Si requirements of coccolithophores, its role in the calcification process, and the impacts of Si availability on the biogeography of these important bloom forming phytoplankton. The results are expected to strengthen our ability to predict the responses of coccolithophores to short and long-term environmental change, and therefore the consequences for the marine biogeochemical cycles in which they participate. In addition to the scientific outcomes, the project provides independent research opportunities to a diverse pool of undergraduate students, provide interdisciplinary training for graduate students, and facilitate the professional development of post-doctoral researchers. Public engagement in the research is facilitated through participant involvement in regional science festivals, public outreach events, production of educational resources, and targeted K-12 summer camp activities.
Calcification in coccolithophores appears to represent a distinct process from silicification in diatoms, another major group of biomineralized phytoplankton. The apparent absence of a requirement for silicon (Si) in coccolithophores has been proposed to play a critical role in their ability to out-compete the otherwise dominant diatoms in areas of low dissolved Si availability. However, the investigators recently demonstrated that some globally important coccolithophores possess diatom-like Si transporters and exhibit an obligate requirement for Si in the calcification process. This discovery has important implications both for phytoplankton ecology and for the evolution of biomineralization. Using a range of physiological, molecular and computational approaches the project will 1) Establish Si requirements of ecologically important coccolithophore groups; 2) Determine the physiological role of Si in coccolithophores; 3) Determine the evolutionary events leading to the differing requirements for Si in calcification; 4) Examine the ecological distribution of Si-requiring coccolithophores, and 5) Determine the impact of the Si requirement on coccolithophore ecology. This project therefore integrates the molecular identification of genes (Si transporters), the physiological role of these transporters, and ecosystem scale models in order to examine how the requirement for Si influences ecosystem functioning and coccolithophore biogeography. The results of this work provides essential data that describes the cellular mechanisms of calcification and the range of physiological diversity between major coccolithophore lineages. The research also explores a previously unforeseen aspect of phytoplankton ecology; examining how the differing requirements for Si in calcifying coccolithophores may have shaped competitive interactions with other phytoplankton over both contemporary and evolutionary timescales. Overall, the research provides novel insights into physiology, ecology and evolution of coccolithophores, including information on how and why coccoliths are produced, which is currently poorly understood. This information is vital in order to understand how coccolithophores have been influenced by past changes in the Earth's climate, and their potential responses to future oceans.
This is a project jointly funded by the National Science Foundation’s Directorate for Geosciences (NSF/GEO) and the National Environment Research Council (NERC) of the United Kingdom (UK).
NERC Award Abstract:
The oceans cover more than three quarters of the surface of the Earth and tiny algae in our seas are responsible for half of all photosynthesis on our planet. These single celled organisms, known as phytoplankton, form the basis of marine food webs and their activities can have an enormous impact on the geology of our planet. One group of phytoplankton known as the coccolithophores produce a covering of calcium carbonate plates (coccoliths) and can form vast blooms in the oceans. When the coccolithophores die, their coccoliths settle to the ocean floor, leading to the formation of sedimentary rocks, such as chalks and limestones. In many parts of the ocean the low availability of nutrients (such as nitrogen and phosphorus) limits phytoplankton growth. Competition for nutrients plays an important role in determining which phytoplankton species can grow in different environments. One of the most successful phytoplankton groups in modern oceans is the diatoms, which are fast-growing, making it difficult for many other phytoplankton types to compete with them. However, diatoms need lots of dissolved silicon to make their silica cell walls. In some marine environments, the availability of silicon limits the growth of diatoms, allowing other phytoplankton (which do not need silicon) to grow in their place. It is commonly thought that the calcifying coccolithophores have no requirement for silicon. However, we have recently discovered that some important coccolithophore species actually possess silicon transporters that are similar to those used by diatoms. Remarkably, we found that these coccolithophores use silicon to make their calcium carbonate coccoliths. Therefore the processes of silica formation in diatoms and calcite production in coccolithophores, which were previously believed to be distinct processes, show a completely unexpected link. These findings have important implications for the evolution of biomineralisation in phytoplankton and for the competitive interactions between coccolithophores and diatoms. Not all coccolithophores show a requirement for silicon. We found that the species responsible for the massive coccolithophore blooms, Emiliania huxleyi, does not possess silicon transporters and exhibits no need for silicon in the calcification process. The absence of a requirement for silicon may have enabled bloom-forming species to grow better in areas where silicon is low (e.g. after a diatom bloom). There is therefore a clear need to understand the role of silicon in coccolithophore biology. In this proposal we will address this issue using a combination of laboratory experiments and computational modelling approaches. Firstly, we will use molecular genetic and laboratory experiments to determine which of the major coccolithophore species exhibit a requirement for silicon. We will then select species for detailed physiological analysis, to determine how silicon contributes to the formation of coccoliths and how coccolithophores take up silicon from the surrounding seawater. These studies will allow us to examine the evolutionary history of the requirement for silicon and determine when certain lineages appear to have lost this trait. Using parameters on Si uptake and usage derived from our experimental work, we will use computer simulations to model global coccolithophore distributions and identify environments where the requirement for Si appears to be playing an important role in coccolithophore ecology. The research will provide novel insight into physiology, ecology and evolution of coccolithophores, including information on how and why coccoliths are produced, which is currently poorly understood. The research will also inform us on the evolution of coccolith formation, which will be vitally important if we are to understand how coccolithophores have been influenced by past changes in the Earth's climate and how they may respond to changes in the future.
| Funding Source | Award |
|---|---|
| NSF Division of Ocean Sciences (NSF OCE) |
[ table of contents | back to top ]